噬菌體
1915年英國人陶爾德(Twort)在培養葡萄球菌時,發現菌落上出現透明斑。用接種針接觸透明斑後,再向另一菌落上接觸,不久接觸的部分又出現透明斑。1917年在法國巴黎巴斯德研究所工作的加拿大籍微生物學家第赫蘭爾(d′Herelle)也觀察到痢疾桿菌的新鮮液體培養物能被加入的某種污水的無細菌濾液所溶解,混濁的培養物變清。若將此澄清液再行過濾,並加到另一敏感菌株的新鮮培養物中,結果同樣變清。以上現象被稱為陶爾德—第赫蘭爾(Twort—d'Herelle)現象。第赫蘭爾將該溶菌因子命名為噬菌體(bacteriophage,phage)。
噬菌體具有病毒的一般特性,是原核微生物的病毒,包括噬細菌體(bacteriophage)、噬放線菌體(actinophage)和噬藍細菌體(cyanophage)。
 偶數噬菌體
偶數噬菌體一、噬菌體的形態結構
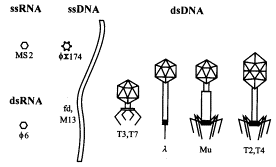
在電子顯微鏡下觀察噬菌體有三種基本形態:蝌蚪形、微球形和絲狀,從結構來看又可分為六種不同的類型,見圖3.5。
圖3.5噬菌體的基本形態和大小
圖中所列的T-系噬菌體是目前研究得最廣泛而又較深入的細菌噬菌體,並對它們進行了從T1-T7的編號,這類噬菌體呈蝌蚪形。
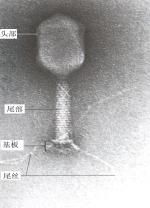
大腸桿菌T4噬菌體為典型的蝌蚪形噬菌體,由頭部和尾部組成。頭部為由蛋白質殼體組成的廿面體,內含DNA。尾部則由不同於頭部的蛋白質組成,其外包圍有可收縮的尾鞘,中間為一空髓,即尾髓。有的噬菌體的尾部還有頸環、尾絲、基板和尾刺(圖3.6)。
 偶數噬菌體
偶數噬菌體圖3.6大腸桿菌(E.coli)噬菌體結構(引自Prescottetal.,2002)和電鏡照片
 偶數噬菌體
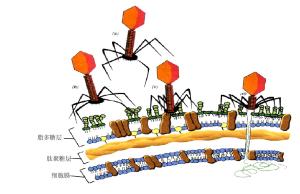
偶數噬菌體圖3.7噬菌體T4吸附在大腸桿菌細胞(Madiganetal.,1997)
A.未吸附B.C.尾部附著D.尾鞘收縮,注入DNA
根據噬菌體與宿主細胞的關係可分為烈性噬菌體(virulentphage)和溫和性噬菌體(temperatephage)。凡侵入細胞後,進行營養繁殖,導致細胞裂解的噬菌體稱烈性噬菌體。而侵入細胞後,與宿主細胞DNA同步複製,並隨著宿主細胞的生長繁殖而傳下去,一般情況下不引起宿主細胞裂解的噬菌體,稱溫和性噬菌體。但在偶爾的情況下,如遇到環境誘變物甚至在無外源誘變物情況下可自發地具有產生成熟噬菌體的能力。
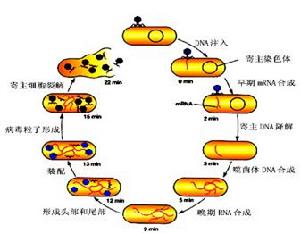
大腸桿菌T系偶數噬菌體的生活周期研究得最早和較深入,這裡以T-系偶數噬菌體為模式介紹噬菌體的增殖,其增殖周期可分為五個階段。
1、吸附(adsorption)
噬菌體侵染寄主細胞的第一步為吸附。吸附過程一方面決定於細胞表面受點的結構,另一方面也取決於噬菌體吸附器官—尾部吸附點的結構。當噬菌體和敏感細胞混合時,發生碰撞接觸,敏感的細菌細胞表面具有噬菌體吸附的特異性受點,噬菌體的吸附點與細菌的接受點可以互補結合,這是一種不可逆的特異性反應。一種細菌可以被多種噬菌體感染,這是因為宿主細胞表面對各種噬菌體有不同的吸附受點。現已證實,大腸桿菌細胞壁的脂蛋白層為T2和T6噬菌體的吸附受點,脂多糖層為T3、T4、T7的吸附受點,而T5噬菌體的吸附受點則為脂多糖—脂蛋白的複合物。吸附時,噬菌體尾部末端尾絲散開,固著於特異性的受點,隨之尾刺和基板固定在受點上。不同的噬菌體粒子吸附於宿主細胞的部位也不一樣,如大腸桿菌T-系噬菌體大多吸附於宿主細胞壁上(圖3.6);大腸桿菌絲狀噬菌體M13隻吸附在大腸桿菌性傘毛的末端;而枯草桿菌噬菌體PBS2則吸附在細菌鞭毛上。至於每個宿主究竟能被多少噬菌體吸附,據測定一般在250~360個即達到飽和狀態,這稱作最大吸附量。
吸附過程也受環境因子的影響,如pH、溫度、陽離子濃度等都會影響到吸附的速度。
侵入即注入核酸。大腸桿菌(E.coli)T4噬菌體以其尾部吸附到敏感菌表面後,將尾絲展開並固著於細胞上。尾部的酶水解細胞壁的肽聚糖,使細胞壁產生一小孔,然後尾鞘收縮,將頭部的核酸通過中空的尾髓壓入細胞內,而蛋白質外殼則留在細胞外。大腸桿菌T-系噬菌體只需幾十秒鐘就可以完成這個過程,但受環境條件的影響。通常一種細菌可以受到幾種噬菌體的吸附,但細菌只允許一種噬菌體侵入,如有兩種噬菌體吸附時,首先進入細菌細胞的噬菌體可以排斥或抑制第二者入內。即使侵入了,也不能增殖而逐漸消解。
尾鞘並非噬菌體侵入所必不可少的。有些噬菌體沒有尾鞘,也不收縮,仍能將核酸注入細胞。但尾鞘的收縮可明顯提高噬菌體核酸注入的速率。如T2噬菌體的核酸注入速率就比M13的快100倍左右。
這個步驟主要指噬菌體DNA複製和蛋白質外殼的合成。噬菌體DNA進入宿主細胞後,立即以噬菌體DNA為模板,利用細菌原有的RNA合成酶來合成早期mRNA,由早期mRNA翻譯成早期蛋白質。這些早期蛋白質主要是病毒複製所需要的酶及抑制細胞代謝的調節蛋白質。在這些酶的催化下,以親代DNA為模板,半保留複製的方式,複製出子代的DNA。在DNA開始複製以後轉錄的mRNA稱為晚期mRNA,再由晚期mRNA翻譯成晚期蛋白質。這些晚期蛋白質主要組成噬菌體外殼的結構蛋白質,如頭部蛋白質、尾部蛋白質等。在這時期,細胞內看不到噬菌體粒子,稱為潛伏期(latentperiod)。潛伏期是指噬菌體吸附在宿主細胞至宿主細胞裂解,釋放噬菌體的最短時間。
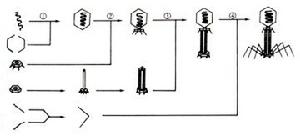
當噬菌體的核酸、蛋白質分別合成後即裝配成成熟的、有侵染力的噬菌體粒子。例如大腸桿菌T4噬菌體的DNA、頭部蛋白質亞單位、尾鞘、尾髓、基板、尾絲等部件合成後,DNA收縮聚集,被頭部外殼蛋白質包圍,形成廿面體的噬菌體頭部。尾部部件也裝配起來,再與頭部連線,最後裝配完畢,成為新的子代噬菌體(圖3.8)。
成熟的噬菌體粒子,除M13等少數噬菌體外,均借宿主細胞裂解而釋放。細菌裂解導致一種肉眼可見的液體培養物由混濁變清或固體培養物出現噬菌斑。絲狀噬菌體fd成熟後並不破壞細胞壁,而是從宿主細胞中鑽出來,細菌細胞仍可繼續生長。大腸桿菌T系偶數噬菌體從吸附到粒子成熟釋放大約需15~30分鐘。釋放出的新的子代噬菌體粒子在適宜條件下便能重複上述過程。
圖3.8T偶數噬菌體裝配過程模式圖
 偶數噬菌體
偶數噬菌體圖3.9噬菌體T4感染各時段的情況(引自Prescottetal.,2002)
 偶數噬菌體
偶數噬菌體三、噬菌體的一步生長曲線
利用烈性噬菌體的生活周期,可以在實驗室條件下獲得噬菌體的生長曲線。這種用來測定噬菌體侵染和成熟病毒體釋放的時間間隔,並用以估計每個被侵染的細胞釋放出來的新的噬菌體粒子數量的生長曲線,稱為一步生長曲線(one-stepgrowthcurve)。它可反映每種噬菌體的三個最重要的特性參數—潛伏期、裂解期和裂解量(burstsize),故十分重要。
將高濃度的敏感菌培養物與適量相應的噬菌體懸液相混,使得一個病毒體與10~100個細菌細胞相混合,這樣的比例係數可降低幾個噬菌體同時侵染單個細菌細胞的機率。經過短時間的培養使噬菌體吸附在細菌上,再用抗病毒血清或離心或稀釋除去未吸附的噬菌體。接著,用新鮮培養液把經過上述處理的細菌懸液高倍稀釋,以免發生第二次吸附和感染。培養後,定時取樣,將含有噬菌體的樣品與敏感細菌培養物混合培養,計算每個樣品在培養基平板表面產生的噬菌斑數目。以培養時間為橫坐標,噬菌斑數為縱坐標,可以繪出一步生長曲線。
如圖3.10所示,噬菌體在吸附和侵入寄主後,細胞內只出現噬菌體的核酸和蛋白質,還沒有釋放出噬菌體,這段時間稱為隱晦期(eclipsephase)。隱晦期間如人為地(用氯仿)裂解細胞,裂解液無侵染性。人們將噬菌體吸附寄主細胞開始到細胞釋放新的噬菌體為止的這段時間稱為潛伏期(latentperiod)。潛伏期過後,噬菌斑數突然迅速上升,表明被感染的細菌已經越來越多地裂解,直至所有感染細胞都被裂解為止,這個時間稱為上升期(riseperiod)。接著,噬菌斑數達到大致恆定,曲線平穩。這個時期即使存在一些未感染的細菌,由於細菌懸液的稀釋倍數很高,使得新釋放的噬菌體不能吸附未感染的細菌。每個感染細菌所釋放的新噬菌體的平均數稱為裂解量。
裂解量=裂解期平均噬菌斑數
潛伏期平均噬菌斑數。
圖3.10噬菌體T2的一步生長曲線A.表示一群噬菌體作用於一群細菌細胞的結果;
B.表示單個噬菌體在一個細菌細胞內的增殖情況
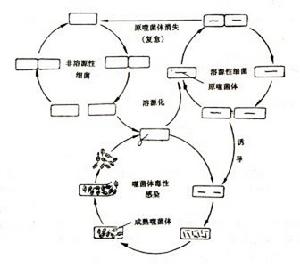
四、溫和性噬菌體的溶源性
溫和性噬菌體侵染宿主細胞後,其DNA可以整合到宿主細胞的DNA上,並與宿主細胞染色體DNA同步複製,但不合成自己的蛋白質殼體,因此宿主細胞不裂解而能繼續生長繁殖。大腸桿菌λ噬菌體就是屬於溫和性噬菌體。整合在宿主細胞染色體DNA上的溫和噬菌體的基因稱為原噬菌體(prophage)。個別噬菌體如大腸桿菌噬菌體P1,其溫和噬菌體的核酸並不整合在細菌的DNA上,而隨著在細胞質膜的某一位點上,呈質粒狀態存在。人們把含有原噬菌體的細菌細胞稱為溶源性細胞(lysogeniccell),並把溫和噬菌體侵入宿主細胞後所產生的這些特性稱為溶源性。
溫和性噬菌體與烈性噬菌體在遺傳上不同。溫和性噬菌體的基因組能整合到細菌染色體中,有一個與細菌染色體相附著的位點,並在其某種基因產物如整合酶的作用下,兩者在此位點發生一次特異性重組。另外,溫和性噬菌體有能編碼合成一種稱為阻遏體蛋白的基因C1,這種阻遏體能阻止噬菌體所有有關增殖基因的表達,從而使其不能進入營養狀態。此外還另有一些基因調節、控制阻遏體的合成,以維持穩定的溶原狀態。如果阻遏體的活性水平減低,不足以維持溶原狀態,原噬菌體就可離開染色體進入增殖周期,並引起宿主細胞裂解,這種現象稱為溶源性細菌的自發裂解,也就是說,極少數溶源性細菌中的溫和噬菌體變成了烈性噬菌體。這種自發裂解的頻率很低,例如大腸桿菌溶源性品系的自發裂解頻率為10-2~10-5。
溶源細胞的誘發裂解用某些適量理化因子,如紫外線或各種射線,化學藥物中的誘變劑、致畸劑、致癌物或抗癌物、絲裂黴素C等處理溶源性細菌,都能誘發溶源細胞大量裂解,釋放出噬菌體的粒子。
溶源細胞的免疫性阻遏體蛋白除阻遏原噬菌體的基因組外,也同樣能阻遏進入溶源菌的其他同型噬菌體的基因組,使其不能在該細胞內複製,因此溶源性菌對同型噬菌體呈現一種特異的免疫現象。例如,含有λ原噬菌體的溶源性細胞,對於λ噬菌體的毒性突變株有免疫性。即毒性突變株對非溶源性宿主細胞有毒性,對溶源性宿主細胞(含λ噬菌體DNA)卻沒有毒性。
溶源性細菌的復愈溶源性細菌有時消失了其中的原噬菌體,變成了非溶源細胞,這時既不發生自發裂解,也不發生誘發裂解,稱為溶源細胞的復愈或非溶源化,這樣的菌株稱為復愈菌株(curingstrain)。
溶源性轉換(Lysogenicconversion)溶源性細菌除具有產生噬菌體的潛力和對相關噬菌體的免疫性外,有時還同時伴有某些其他性狀的改變,這種其他性狀的改變稱為溶源性轉換。例如白喉棒狀桿菌產生白喉毒素是因為原噬菌體帶有毒素蛋白的結構基因;肉毒梭菌的毒素、金黃色葡萄球菌某些溶血素、激酶的產生都與溶源性有關;沙門氏菌、痢疾桿菌等抗原結構和血清型別也與溶源性有關。現在知道越來越多菌類的各種性狀都受到溶源性的影響。這種現象很象腫瘤病毒能使正常細胞轉化為腫瘤細胞的轉化現象。噬菌體與宿主菌細胞之間的關係如圖3.11所示。
 偶數噬菌體
偶數噬菌體