
三角渦蟲
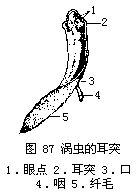 三角渦蟲外形
三角渦蟲外形外形特徵
渦蟲體柔軟,扁平葉狀,背面稍凸,多褐色,腹面色淺,前端頭部呈三角形(故得名),兩側各有一發達的耳突(auricle),司觸覺和嗅覺,頭部背面有兩個黑色眼點(eyespots),可感覺光線的明暗,口位於腹面近體面後1/3處,稍後方為生殖孔,無 肛門,身體腹面密生纖毛,與渦蟲運動有關。
結構與機能
(1)體壁與運動 渦蟲的體壁是典型的皮肌囊。表皮(epidermis)由外胚層來的柱狀上皮細胞組成,其間有大量的腺細胞、桿狀體(rhabdites)。桿狀體是由上皮的一種腺細胞(成桿狀體細胞或稱桿狀體腺細胞rhabdite gland cell)分泌的,細胞體常沉入表皮之
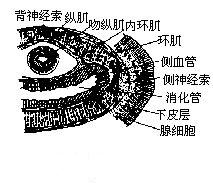 三角渦蟲的橫切面
三角渦蟲的橫切面下的實質中。其他腺細胞成熟時也常沉入於實質,僅其“頸部”留在表皮,他們分泌的黏液有多種功能(如保護蟲體免於乾燥,利於氣體交換,有助於運動等)。桿狀體是渦蟲的一種分泌物。為一種特殊分層的超微結構。當渦蟲受到刺激時排出體表,遇水常彌散出有毒性的黏液,以捕食和防禦敵害。腹面的表皮有纖毛,纖毛之間有短的微絨毛。表皮之下為非細胞結構有彈性的基膜(basal lamina 或 basal membrane),其下面是中胚層形成的肌肉層,由外向內分為三層,一次為環肌、斜肌和縱肌。渦蟲主要依纖毛擺動,藉助於其分泌的黏液,滑動於物體表面,由於渦蟲具有發達、複雜的肌肉系統,能使其做出不同的運動,如爬行蠕動、收縮伸展、扭動、波動、翻轉和翻跟頭等。
(2)實質與其潛能 實質(parenchyma)來源於中胚層,充滿於體壁和內部器官之間,疏鬆地互相聯繫在一起,可儲存養分和水分。一般說,實質是一種結締組織,有細胞和細胞外間質【纖維和/或液體。僅無腸目渦蟲例外,幾乎無細胞外間質。實質細胞不同,已確定功能的細胞:表皮取代細胞(epidermal replacement cell),緊貼體壁之下,從實質移至體表,取代任何被損傷或破壞的細胞;成新細胞(neoblast,也譯為成年未分化細胞)】,是全能細胞(totipotent cell),它對損傷癒合和再生很重要,也可產生表皮取代細胞;另一種為固定實質細胞(fixed parenchyma cell),是大的分支細胞,它與其他實質細胞以及表皮和胃層細胞構成間隙連線(gap junction)。這使蟲體所有組織層連線在一起。間隙連線時代謝物低阻力運輸的細胞間通道。而且它們的存在表明,由它們連線的細胞網狀結構,在缺乏循環系統的扁蟲,或許是一種特殊的細胞間運輸系統。此外,有些渦蟲有實質的色素細胞(pigment cell)和載色體(chromatophore)。後者在細胞中的色素集中或分散時,能使細胞變深或變淺。載色體受腦的控制。
(3)攝食和消化 消化系統 口在腹中線後1/3處,口後為咽,在發生中管狀咽進一步向內翻折成咽鞘及肌肉質的咽,兩者之間的空間稱之為咽囊或咽腔,這種咽
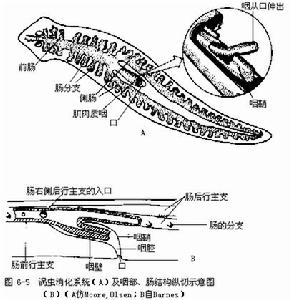 三角渦蟲消化系統及咽部、腸結構縱切示意圖
三角渦蟲消化系統及咽部、腸結構縱切示意圖稱為摺疊咽(plicate pharynx)。咽可從口中伸出或縮回。咽後為腸,腸壁來自內胚層,為一單層柱狀上皮細胞,其中有大量的腺細胞和吞噬細胞。腸分3支主幹,一支向前,兩支向後,分別位於咽囊兩側,每支主幹又反覆分出末端為盲端的小支,無肛門,不能消化的食物仍由口排出。
已知渦蟲的取食行為是由食物放出的物質誘發的。取食時蟲體先後分泌黏液,黏纏固定捕獲物,藉助咽腺分泌蛋白水解酶將咽管插入捕獲物體內,吮其內組織液體,或將食物進行部分消化,然後吸入腸內,由腸腺細胞分泌內肽酶(endopeptidase)先行細胞外消化,消化後的碎片由吞噬細胞吞噬,並在小囊泡中由內肽酶低PH下開始細胞外消化。吞噬作用後8~12h,小囊泡變成鹼性,它標誌外肽酶(exopeptidase)、脂酶(lipase)和糖酶(carbohydrases)的出現,進行完全消化。由於腸分支較多,無疑增加了消化吸收面積,也運輸營養到身體各部分。可見其作用仍屬胃循環系。
(4)呼吸與循環 渦蟲無特殊的呼吸、循環器官,依靠體表擴散作用進行氣體交換,借網狀的是指細胞增加表面面積,由其中的液體運送和擴散新陳代謝的產物。
(5)排泄與滲透調節 排泄系統為原腎管型,由焰細胞和排泄管組成。在蟲體兩側有一對彎曲、多次
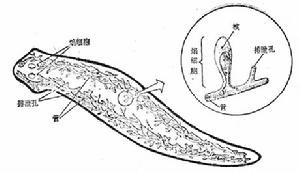 三角渦蟲的排泄系統
三角渦蟲的排泄系統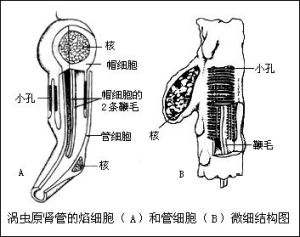 三角渦蟲原腎管的焰細胞和管細胞微細結構圖
三角渦蟲原腎管的焰細胞和管細胞微細結構圖分支的總行排泄管,每一小分支細管的末端連著焰細胞(即帽細胞和管細胞)。通過焰細胞收集體內多餘的水分和液體廢物,經排泄管由體背面的排泄孔排出體外。原腎管的主要功能是調節體內水分的滲透壓,時也排除一些代謝廢物。
(6)神經與感官 梯形神經系統,頭部有一對腦神經節,由此分出一對腹神經索通向體後,在腹神經索之間有橫神經相連,因而構成梯形。渦蟲背部的一對眼點是由色素細胞和視覺細胞所構成,它們只能辨別光線的明暗,不能看物象;
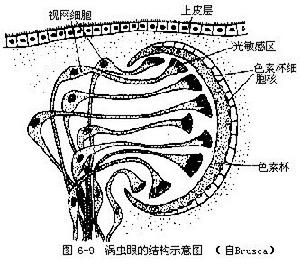 三角渦蟲的眼的結構示意圖
三角渦蟲的眼的結構示意圖耳突在頭的兩側,有許多感覺細胞,司觸覺和嗅覺,在表皮內還分布著許多觸覺細胞,渦蟲對食物是正向反應,對光線的反應是避強光,尋找暗的微光,夜間活動強於白晝。
 三角渦蟲的神經系統
三角渦蟲的神經系統生殖與發育
三角渦蟲具有性生殖和無性生殖兩種方式。有性生殖雌雄同體,雌雄系統相當複雜。
雄性生殖系統 在體之兩側有很多精巢,每一精巢有一輸出管(vas efferens),匯合在兩側各成一輸精管(vas deferens),到身體中部膨大,一般稱為貯精囊(seminal vesicle),兩貯精囊匯入多肌肉的陰莖(penis),在陰莖基部有很多單細胞腺體稱為前列腺(prostate glands),開口於生殖腔(genital atrium)。
雌性生殖系統 在身體前方兩側各有一卵巢,每一卵巢有一輸卵管(oviduct)向後行,同時收集由卵黃腺(vitellaria)來的卵黃,兩條輸卵管在後端匯合形成陰道(vajina,即為雌生殖腔),通入生殖腔中,由陰道前端向前伸出一條受精囊(seminal receptacle,也成交配囊,copulatory bursa)在交配時接受和儲存對方的精子。渦蟲雖為雌雄異體,但需要交配進行異體受精(cross-fertilization)。
渦蟲交配時,兩蟲各翹起尾端的一段,腹面配合,各從生殖孔內伸出陰莖進入對方的生殖腔內,輸入精子,行體內受精,然後兩蟲分開。對方的精子暫時儲存在受精囊內,當卵巢排卵時,從囊內游出,沿陰道、輸精管到達輸卵管前段與卵受精。受精卵附以卵黃腺所產的卵黃細胞移至生殖腔,幾個受精卵和不少卵黃細胞一起被生殖腔分泌的黏液(形成皮膜)裹住,成為卵囊(egg capsule)或卵袋(cocoon),最後從生殖孔排出。三角渦蟲的卵袋是圓球形,有一小柄附於浸
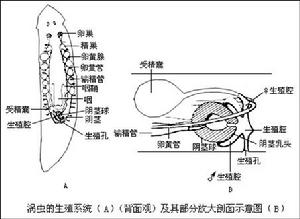 三角渦蟲的生殖系統及其部分放大剖面示意圖
三角渦蟲的生殖系統及其部分放大剖面示意圖在水中的石塊或其他物體上。夏季產生的受精卵,卵袋較薄,幾天后即可孵化;秋季產生的受精卵,卵袋較厚,為休眠卵,翌春孵化。直接發育(淡水或陸地生活者),幼蟲孵出後就吸食卵袋中的卵黃,然後離開卵袋,發育成為成蟲。
三角渦蟲進行有性生殖外,尚可進行無性生殖,以分裂方式進行無性生殖。分裂時以蟲體後端黏於底物上,蟲體前端繼續向前移動,直至蟲體斷裂成兩半。其分列面常發生在咽後,然後各自再生出失去的一半,形成兩個新個體。
再生
三角渦蟲的再生能力很強,若將它橫切為兩段,每一段都會再長出其失去的一半,成為一條完整的渦蟲,甚至分割為許多段時每一段也能再生成一個完整的渦蟲。還能進行切割或移植,產生二頭或二尾的三角渦蟲。三角渦蟲的再生表現出明顯的極性,再生的速率由前向後呈梯度遞減,即前端生長發育最快,後端最慢。現代研究的中心,在於回答兩個問題:(1)是什麼控制結果類型的重建?(2)再生細胞的來源是什麼?目前看來,實質中的成新細胞(neoblast)是全能的、未分化細胞,它可構成胚基(blastema)來源的細胞。成新細胞相當於海綿的原細胞(archeocyte)和腔腸動物的間細胞(interstitial cell)。但也有人認為,胚基未分化的細胞可從已分化的細胞產生,如肌細胞通過去分化或細胞反轉到全能的胚胎未分化狀態。現在正研究搜尋尋找控制再生的機制。例如,生長因子(growth factor)可能開始再生。HOX基因控制前後端極性,並可表明前後端、背腹極性的特殊區域標誌(蛋白質)。一旦這些控制因子被鑑別出來,有希望通過刺激再生作用,用於治療人類脊髓損傷等疾病。
此外,當三角渦蟲飢餓時,內部的器官(如生殖系統等)逐漸被吸收消耗,唯獨神經系統不受影響,一旦獲得食物後,各器官又可重新恢復,變成正常的體型,這也是再生的一種方式。
